|
The Natural History of Tarantula
Spiders Introduction Taxonomic Position
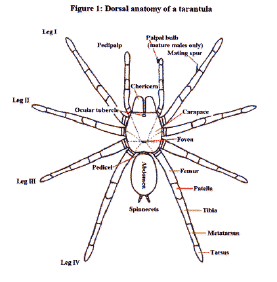
The theraphosids share the mygalomorph infraorder with 14 other spider families. They differ from these other families in a number of ways, but no single feature can used to distinguish them. Typically tarantulas possess thick scopulae (pads of specialised hair) on the undersides of their metatarsi and tarsi. Their tarsi also have claw tufts, which as their name suggests, are tufts of scopulae located next to their tarsal claws. The vast majority of tarantulas have long finger-like end segments to their posterior spinnerets. An additional differentiating feature is the possession of a prominent anterior lobe on the maxilla. This feature is useful in separating the Theraphosidae from the very similar Barychelidae, which have a weak anterior lobe. The Theraphosidae family is further subdivided into 13 subfamilies. The actual number of subfamilies varies depending on which taxonomist you talk to. This is because not all agree on the validity of some of the taxa. Of the New World subfamilies the largest by far is the Theraphosinae. This includes the majority of terrestrial species found in the Americas (e.g. Grammostola, Aphonopelma, Theraphosa and Brachypelma). The Aviculariinae subfamily comes second with three of the four genera being arboreal in nature. This subfamily houses the genera Avicularia, Ephebopus, Pachistopelma and Tapinauchenius. Some taxonomists feel that Psalmopoeus should also be included in this subfamily, but the evidence for this is weak to say the least. Most taxonomists believe that Psalmopoeus belongs to the Selenocosmiinae subfamily. The cave dwelling members of the genus Spelopelma are given their own subfamily (Spelopelminae). Likewise the genus Acanthopelma is also in a monogeneric subfamily (Acanthopelminae). Four of the New World genera are referable to a temporary group termed ‘New World Ischnocolinae’. These are not closely related to the Old World Ischnocolinae. Africa is the exclusive home of the Eumenophorinae, Stromatopelminae and Harpactirinae subfamilies. The most familiar members of the Eumenophorinae are the Hysterocrates spp. and Citharischius crawshayi. Popular captive genera of the Harpactirinae include Pterinochilus and Ceratogyrus. Members of the Ischnocolinae and Selenogyrinae subfamilies are also represented in the African theraphosid fauna. Asia also plays host to its fair share of endemic subfamilies (Ornithoctoninae, Thrigmopoeinae and Poecilotheriinae). Although one genera from the Selenocosmiinae exists in the New World, the primary home of this subfamily is South East Asia. Theraphosids from the Selenogyrinae and Ischnocolinae taxa are also found in the Asiatic region. The only subfamily found in Australia is Selenocosmiinae. Like Australia, mainland Europe also has an impoverished tarantula fauna. Its sole representative genus being Ischnocolus (subfamily Ischnocolinae). External Anatomy (Figures 1
& 2)
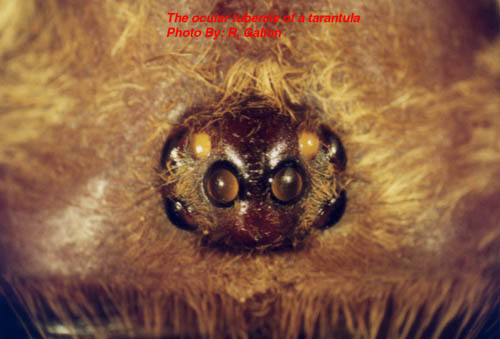
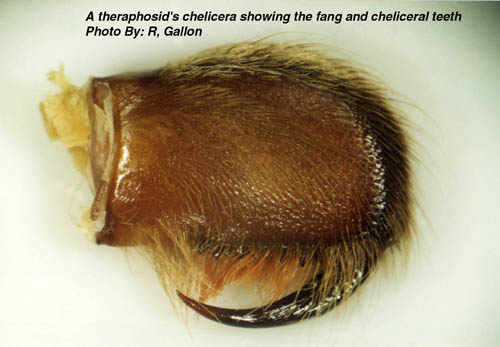
Most theraphosids have eight eyes, but some members of a Mexican cave-dwelling genus (Spelopelma) are totally blind (i.e. have lost all their eyes). Immediately in front of the carapace is a pair of short, thick appendages called chelicerae. On their ventral surfaces each chelicera is armed with a venomous fang. The fangs, in their relaxed state, fold next to a row of spikes or teeth (cheliceral teeth)
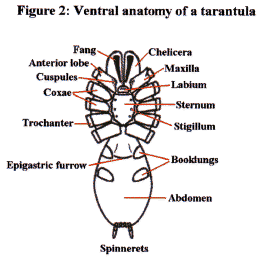
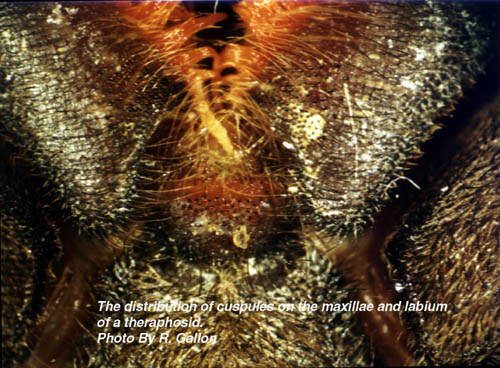
The body and legs of all tarantulas are distinctly hairy. Most of the visible hair (setae) acts as waterproofing and as a barrier to parasites. There are however extremely fine setae present on the legs and palpi which are sensitive to air movements (trichobothria). These trichobothria aid in prey/predator detection for the essentially nocturnal tarantula. Other specialised setae are also present on the legs, which are used to ‘taste’ their environment. The underside of the cephalothorax is furnished with a central shield (sternum) surrounded by the coxae of the palpi and legs. The sternum possesses three pairs of bald spots known as stigilla, which are sites of internal muscle attachment. Connected to the front of the sternum is a smaller plate called the labium (covers the mouth). The labium is covered with small, sclerotised (hardened) cuspules.
The second of the two body segments is the abdomen or opisthosoma. This is attached to the cephalothorax by a narrow waist (pedicel). The abdomen, unlike the rest of the body, is able to expand and contract to accommodate food and developing eggs. In most New World species a proportion of the dorsal pelage is specialised into defensive urticating setae. Two pairs of spinnerets, from which silk is spun, can be found at the end of the abdomen
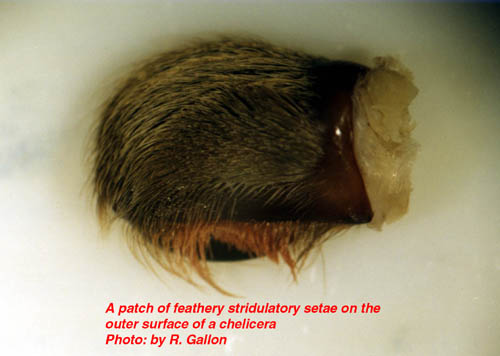
Two pairs of pale booklungs can be seen on the underside of the abdomen. These internal structures, as their name suggests, are responsible for gaseous exchange (oxygenating and removing carbon dioxide from the blood or haemolymph). The openings to the booklungs can be discerned as slits at the rear of the pale regions. Located between the front pair of booklungs is the transverse genital opening or epigastric furrow. In females this is a wide opening, but very much narrower in the males. Distribution and Life Style Most tarantulas adopt one of three life styles thus:- The vast majority frequent burrows, which are usually, self constructed, but sometimes adapted from abandoned mammal or reptile burrows. All species found in the more northerly and southern ranges frequent burrows. Typically their burrows consist of a simple tube sunk into the earth at an angle terminating with an ovoid living chamber. In some species the burrow system can be more elaborate with additional chambers and entrance tubes (e.g. Ischnocolus spp.). Some species embellish the entrance to their burrow with a turret composed of silk, plant material and soil (Haplopelma spp., Lampropelma spp. and Ephebopus spp.). This turret prevents surface water from flooding the burrow. It is believed that some species do not construct permanent burrows. These are sometimes referred to as vagabond species. They wander about during the night taking refuge by day under whatever shelter they come across. It has been suggested that Cyriocosmus ritae and Lasiodora parahybana behave in this fashion, but as yet this life strategy is not widely excepted and further field work will be required to verify such behaviours. The third life style is that of living in trees (arboreal). This behaviour is less common than the terrestrial mode of life but nonetheless six theraphosid subfamilies have members which occupy this niche. South America has the greatest number of arboreals with two representative subfamilies [Aviculariinae (Avicularia, Tapinauchenius, Pachistopelma) and Selenocosmiinae (Psalmopoeus)]. Africa has another two subfamilies with arboreal representatives [Harpactirinae (some Pterinochilus) and Stromatopelminae (Heteroscodra, Stromatopelma)]. Asia also possesses arboreals contained in two of its subfamilies [Ornithoctoninae (Phormingochilus) and Poecilotheriinae (Poecilotheria)]. The arboreals either live in specially constructed silken tube webs (e.g. Avicularia avicularia), rot holes, behind loose bark,or amongst epiphytic plant growth. Usually tarantulas live a solitary existence with one spider occupying one retreat. There are however some examples where more than one spider cohabit. The most famous example of this exists amongst some of the members of the genus Poecilotheria. In the wild specimens of P. subfusca in varying sizes have been found sharing the same tree hollow. In captivity specimens of P. regalis have been successfully maintained together in single vivaria. Many reasons for this behaviour have been put forward but perhaps the most likely explanation is that suitable retreats are at a premium which necessitates this sharing. Some African species also display some degree of retreat sharing. Females of a Hysterocrates sp. from the island of Sao Tom� have been found sharing their burrow with large immature specimens (presumably her progeny). The large immatures even hunt from their mother’s burrow. From captive observations the majority of theraphosid spiderlings disperse en mass from the female’s retreat at the 1st instar stage. Spiderling/mother group feeding has also been witnessed in captive specimens of a Hysterocrates species originating from Cameroon. Burrow Inhabitants Recently a small group of flies have been found cohabiting with tarantulas in a quite intimate way. These flies belong to the genus Megaselia within the family Phoridae. The adults are frequently found in close association with certain tarantulas. These flies lay their eggs on the spider or on some of the detritus within the burrow. The maggots then consume prey remains and even move to the spider’s mouth to feed on the spider’s meal. Adult flies are also believed to drink the fluids exuded by the tarantula whilst it is moulting. Of the three tarantula associating species found so far all have proved host specific (i.e. one fly species per tarantula species). To date Theraphosa blondi, Megaphobema robustum and Pamphobeteus vespertinius are known to play host to these intriguing flies. Parasites Prey Theraphosids are equipped with two large fangs supplied with venom from glands housed in their chelicerae. Tarantula venom is not considered to be of medical importance to humans. Of the hundreds of reported cases of tarantula bites very few cause anything more serious than temporary local inflammation. Defence Strategies Two distinct modes of delivery are known. The simplest delivery is practised by Avicularia spp., which press their abdomens' against their enemy’s skin. These type II urticating setae uniquely impale the attacker with a pointed end situated near the hair’s attachment point. These setae are held in the victim's skin by a battery of backward pointing barbs. A more elaborate delivery is practised by the other New World species in the subfamily Theraphosinae. These possess a range of structurally different, barbed, urticating setae that they are able to brush into the air with their rear legs (resulting in a bald abdominal patch). These airborne setae penetrate the enemy, and are then able to work their way deeper into the attacker’s skin by virtue of a series of backward pointing barbs. Each type of urticating setae is believed to target different enemies, although the targets for some types are still unknown. Types III and IV are the most irritant to mammalian predators. Type III setae may also target invertebrates. Some species incorporate urticating setae into their silken retreats and egg sacs (Avicularia spp. and Theraphosa blondi respectively). This is thought to discourage dipteran maggots from consuming their eggs and young. To date no evidence has been found to suggest that urticating setae possess any chemical irritants. Their irritant effects are believed to be solely due their physical morphology. Old World species (and Psalmopoeus spp.) do not possess urticating setae. In general these species rely on threat posturing to discourage predation (many New World species also use threat posturing). The threat posture is assumed when the spider rears up on its back two pairs of legs. In this position the spider extends its front two leg pairs and palpi vertically into the air. In some species the chelicerae are also opened ajar. Under continued provocation the spider will lunge forward periodically and a bite may be delivered. The underside of most species is dark in colour and this makes the red setae surrounding the mouth very prominent. Members of the genus Poecilotheria have brightly coloured bands of yellow, white and black under their two front pairs of legs (P. ornata also possesses red under its palpi). These brightly coloured areas are thought to act as threat devices to scare potential predators. Many species are also able to produce threatening sounds by means of specialised stridulatory setae (hooked, feathered, barbed or spiked).
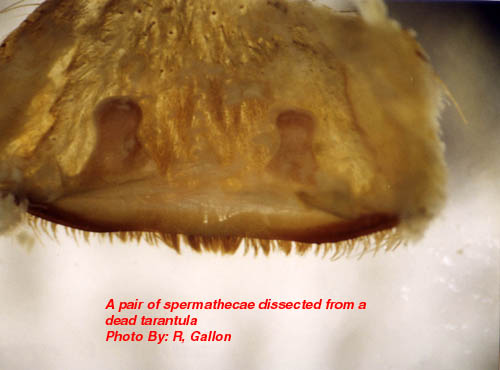
Reproduction and Growth Mature male theraphosids, like other spiders, possess secondary sexual organs at the ends of their pedipalps. These highly sclerotised (hardened) sperm storage vesicles (palpal bulbs) are used to inject sperm into the female’s genitalia. Prior to mating the male tarantula needs to charge these bulbs with sperm. Firstly he spins an angled sheet of silk against a fixed object such as a burrow wall. He then crawls on his back under this lean-too structure and deposits a patch of sperm onto the underside of this web (sperm web). The male then exits the structure and sits on top of it so that his palpi can reach under the sperm web canopy. The male proceeds to dips his palpal bulbs into the patch of sperm, which is draw into these structures. Once completed the male usually destroys the sperm web (at least in captivity) leaving a thin white strand of silk as the only evidence of his activity. The mature male will then leave his retreat under the cover of darkness and set out in search of females. He will cover a lot of ground before picking-up the pheromones of a receptive female, which he will then home in on. Uniquely females of Aphonopelma hentzi construct large silken balloons in the grass above their burrows. These structures are thought to act as pheromone beacons for wandering males. Once the male reaches his target he will proceed to court the female with various species-specific signals. The display often consists of pedipalp drumming on the ground and quivering various parts of the body (e.g. the abdomen in Pterinochilus murinus). In some species the female will also pedipalp drum to signal her receptivness (e.g. Brachypelma albopilosum). The male will stand over the entrance and lure the female from her retreat. On contact there is copious amounts of leg fencing with both spiders waving their legs together. The male will then attempt to gain high ground. Males of many species possess tibial spurs on the ventral surfaces of their front legs’ tibiae. These spurs are used to press against the ventral surface of the female’s chelicerae allowing the male to push the female upwards. Species that do not possess tibial spurs also push females up with their front legs, which may be armed with fringes of hair to aid this (e.g. Stromatopelma spp.). Once the female is secure the male will attempt to extend his pedipalps towards the female’s genital opening. The male will then insert his palpal bulbs and discharge his sperm. This is heralded by a collapse of the female who is pulled towards the male in the process. The male will then remove his palpal bulbs, disengage his front legs and beat a hasty retreat. Once at a safe distance he will clean his bulbs with his fangs and mouth. For a short period of time the female may remain in a trance like state at the scene of the mating. In some species the female may attempt to devour the male following mating (e.g. Brachypelma albopilosum). Smaller or weaker males may fall victim, however larger males will usually fend off the female and survive the encounter. In the vast majority of cases the male will survive and once he has recharged his palpal bulbs he will be able to mate with another female. On maturing male tarantulas live for between two months to three years depending on the species. Bearing in mind these are observations made on captive material, one would only expect wild males to live for a few months at the most. The female tarantula now possesses a batch of sperm in her sperm storage organs or spermathecae. The spermathecae are sclerotised pockets located just under the female’s genital opening
At emergence time the spiderlings may leave the egg sac in one of two stages depending on the species. In Avicularia spp. the young emerge as 1st instar spiderlings which are hairy and fully mobile. Most other species emerge as postembryos (sometime called nymphs) which are pale white, essentially bald, relatively immobile creatures. Postembryos require a few weeks before they darken and moult into 1st instars. It should be noted that Avicularia spp. postembryos moult into 1st instars within the egg sac. After a few more weeks the 1st instars disperse nocturnally en mass from the female’s retreat (except some Poecilotheria spp. and Hysterocrates spp.). The highly mobile spiderlings eventually settle down to construct burrows or arboreal retreats depending on the species. Between one and several years elapse before the spiderlings reach maturity with the males of most species maturing earlier than their sisters. In order for spiders to grow it is first necessary for them to shed their inflexible exoskeleton. Usually the tarantula will spin a mat or hammock of silk on which to moult. Once this is completed the spider usually flips onto its back. Pressure generated from within spits the old exoskeleton under the edge of the carapace. This split then spreads along the sides of the abdomen. Rhythmical movements push the old exoskeleton from the cephalothorax’s appendages. Once the spider’s appendages are free a little hardening time, with additional leg flexing elapses before the spider rights itself. Immediately following moulting the spiders fangs are white and its pelage paler than usual. Over the proceeding days the spider hardens-up and normal, brighter, coloration is assumed. Tarantulas like other spiders are able to replace lost appendages (chelicerae, fangs, palpi, legs and spinnerets) when they moult. These re-growths tend to be smaller than normal, but subsequent moults allow normal proportions to be attained. Immediately before and after moulting the tarantula will be unable to feed and often seals its retreat to prevent disturbance during this delicate time. It is interesting to note that tarantulas also shed some internal organs along with the exoskeleton. These include the linings of the oesophagus, sucking stomach, venom ducts, book lungs and spermathecae. An important factor in the shedding of the spermathecae is the loss of any stored sperm. As a result unmated, freshly moulted females are unable to produce fertile egg sacs. Spiderlings require several moults before they mature. This can take anywhere between 1 and 10 years depending on species, food availability and gender. Female tarantulas like other mygalomorphs are able to moult after reaching maturity. This ability is very rare amongst araneomorphs. This post maturation moulting ability allows the female to grow to large proportions. She may live form anywhere between 6 and 30 years. It is possible that some species may even reach 50 years old, but that is as yet unproven. Male tarantulas on the other hand rarely moult once they are sexually mature. Under captive conditions males of Brachypelma vagans and Grammostola rosea have completed moults following maturity, but these usually result in the loss of the palpal bulbs. Tarantulas are famed as the giants of the spider world. The latest Guinness Book of Records (1999) notes that the largest know spider was a mature male Theraphosa blondi captured in Venezuela. This Herculean spider had a leg-span of 28cm (11 inches). There are reliable reports of a captive male Pseudotheraphosa apophysis with leg-spans in excess of 30cm (12 inches). The heaviest tarantula known to Guinness is an unspecified female from Surinam, which weighted in at 122.2 grams (about 4 ounces). Unconfirmed reports exist that captive, female specimens of T. blondi have tipped the scales at six ounces. Round up Readers wishing to follow-up some of the information presented in this whistle stop article are encouraged to seek out the original source material, most of which is listed in the bibliography that appends the following publication: - Breene R. G. et al. 1996. Tarantulas of Texas, their medical importance. American Tarantula Society, Artesia. Copies of this useful work can be obtained from the American Tarantula Society (refer to the B.T.S. links page for their web site address). Plate 1: An adult female African theraphosid (Pterinochilus sjoestedti) Plate 2: The ocular tubercle of a tarantula. Plate 3: A theraphosid’s chelicera showing the fang and cheliceral teeth. Plate 4: Palpal bulb at the end of a mature male’s left pedipalp. Plate 5: A mating spur on the tibia of leg I from a mature male tarantula. Plate 6: The distribution of cuspules on the maxillae and labium of a theraphosid. Plate 7: The spinnerets of a tarantula viewed from below. Plate 8: A patch of feathery stridulatory setae on the outer face of a chelicera. Plate 9: A pair of spermathecae dissected from a dead tarantula. |